
Мы привыкли представлять себе ДНК в виде двойной спирали — но это лишь одно из множества ее обличий. С тех пор, как Уотсон и Крик опубликовали свою модель, в клетках человека нашли тройную и четверную спираль ДНК, а еще кресты, шпильки и другие варианты переплетения — некоторые проще нарисовать, чем описать словами.
Набросать идей
Уотсон и Крик не были единственными, кто корпел над трехмерной моделью ДНК. Они даже не были первыми. На обрывках биохимических данных можно было построить самые разные молекулярные формы, и вариантов было множество.
Условия задачи у всех были одинаковы. На начало 1953 года уже было понятно, как устроен нуклеотид:
- остаток фосфорной кислоты,
- сахар,
- одно из азотистых оснований: аденин (А), гуанин (Г), тимин (Т) или цитозин (Ц).
Еще было известно, что азотистые основания разбросаны по цепи не случайно: в любой молекуле ДНК суммарное количество аденинов и гуанинов строго равнялось количеству тиминов и цитозинов. Кроме того, на всех рентгеновских снимках Розалинд Франклин и Рэймонда Гослинга, независимо от того, какой участок ДНК на них был запечатлен, сама нить имела одну и ту же толщину. Это означало, что форма остается неизменной при любой последовательности нуклеотидов.
Из этих вводных Лайнус Полинг и Роберт Кори собрали свою модель — тройную спираль, ощетинившуюся со всех сторон азотистыми основаниями (фосфату и сахару биохимики отвели роль внутреннего стержня). Эта конструкция выглядела неустойчивой: было непонятно, почему отрицательно заряженные фосфатные группы в центре спирали не отталкиваются друг от друга.

Структура ДНК по версии Полинга и Кори
Эту проблему решил Брюс Фрезер, вывернув конструкцию наизнанку: в его варианте три нити смотрели фосфатами наружу. Азотистые основания были обращены внутрь, однако Фрезер так и не смог объяснить, по какому принципу они соединены.
Модель Уотсона и Крика с закрученной вправо двойной спиралью оказалась самой устойчивой. Как и Фрезер, ученые расположили фосфаты снаружи, а азотистые основания — внутри. Был в этой модели и четкий принцип их противопоставления: А на одной цепи всегда соединялся с Т на другой, а Г — с Ц. Это объясняло, почему толщина конструкции стабильна — пары А-Т и Г-Ц примерно одинакового размера.
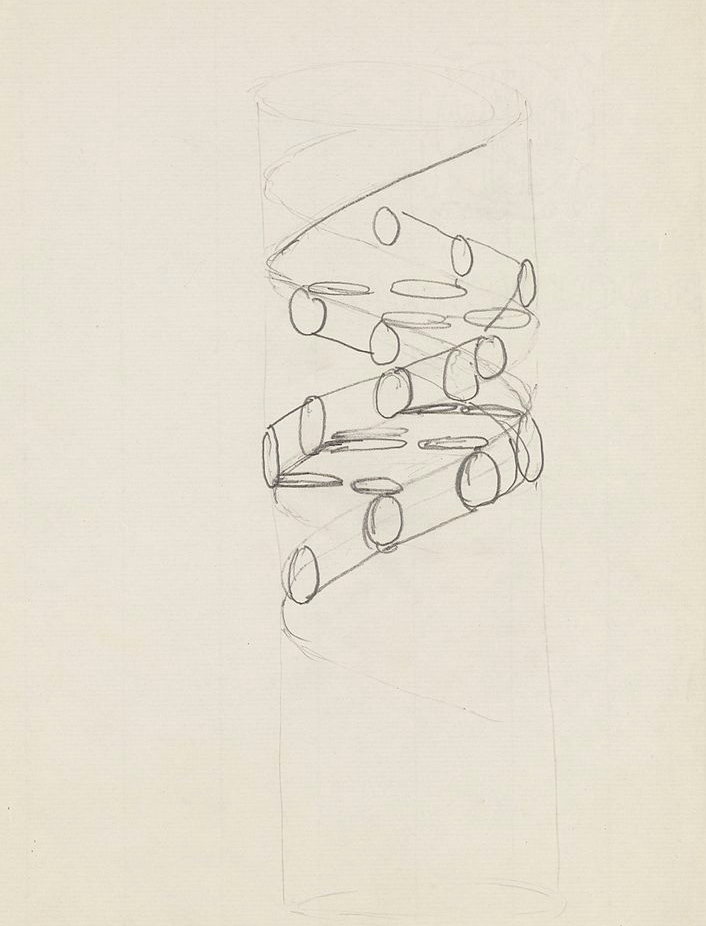
Карандашный набросок структуры ДНК, сделанный Фрэнсисом Криком
Потом были и другие попытки пересобрать ДНК в новую форму. Голландский биохимик Карст Хугстин, например, заметил, что можно соединить те же самые пары нуклеотидов другими гранями, — так спираль тоже оставалась стабильной, но получалась тоньше. Другие авторы изображали ДНК в виде спирали с чередующимися правым и левым поворотами, или даже в виде двух двойных спиралей, которые образуют единую четверку. И хотя существование Уотсон-Криковской двойной спирали с тех пор много раз подтвердилось, в XXI веке продолжают размышлять о том, какие формы принимает нить ДНК внутри клетки, где ее разглядеть намного сложнее, чем в пробирке. Правда, ни одна из альтернативных идей до сих пор не оказалась достаточно хороша, чтобы отказаться от классической правозакрученной двойной спирали.
Уотсон и Крик сделали нечто большее, чем просто разрешили споры о форме ДНК. Их модель сразу же объяснила, как эта форма работает: взаимно однозначное соответствие делает каждую нить шаблоном для другой. Имея только одну из цепей, по ней всегда можно восстановить вторую — на этот принцип опираются все современные модели передачи генетической информации.
Тем не менее, большинство «отвергнутых» идей в чем-то оказались верны. За почти 70 лет пристального разглядывания ДНК в ней удалось обнаружить практически все возможные виды соединения оснований, другие спирали и даже левый поворот.
Свернуть не туда
Уже сама по себе двойная спираль может быть устроена по-разному. Это заметила еще Розалинд Франклин, хотя и не предполагала, что перед ней спираль, да еще и двойная. В обычных условиях, напоминающих внутриклеточные, ДНК на снимках биолога имела «рыхлую» форму, которую Франклин назвала В-ДНК. Но если влажность в пробирке опускалась ниже 75 процентов, получалась А-ДНК, пошире и поплотнее.
А (слева) и В (справа) формы ДНК, какими их увидела Розалинд Франклин
Как выяснилось потом, А-ДНК действительно закручена туже: в ней на виток спирали уходит 10 нуклеотидов, а не 11, как в В-ДНК. И расположены они не перпендикулярно оси симметрии спирали, а под углом: если в В-ДНК нуклеотиды обычно изображают горизонтальными черточками, в А-ДНК их следовало бы рисовать косыми.
Уотсон и Крик выбрали В-ДНК в качестве основы для своей модели и не прогадали. Позже оказалось, что В-вариант действительно встречается в клетке гораздо чаще, и сейчас его считают основной формой существования ДНК, а все отклонения часто обозначают общим термином «не-В ДНК».
Более того, реальная двойная спираль почти никогда не соответствует своей идиллической модели. В живых системах В-ДНК, как правило, скручена чуть сильнее, чем предсказывали Уотсон и Крик, и среднее число нуклеотидов на виток спирали в ней — не 10 и не 11, а около 10,5. Кроме того, отдельные пары нуклеотидов постоянно отклоняются от положенной «горизонтали» (это называют «пропеллерным поворотом») поэтому спираль никогда не бывает абсолютно гладкой и ровной — то тут, то там по ее бокам торчат шероховатости: концы нуклеотидов под разными углами.
«Пропеллерный» поворот нуклеотидов в В-ДНК
Позже оказалось, что витки спирали могут не только лежать туже или расслабленнее, но и вовсе закручиваться против часовой стрелки (например, влево закручена спираль башни «Эволюция» в Москва-сити, явно символизирующая нить ДНК). По странному стечению обстоятельств, именно такую ДНК увидели в 1979 году, когда появилась наконец возможность рассмотреть нуклеиновые кислоты с высоким разрешением. Это все еще была двойная спираль, но совсем другой формы: 12 нуклеотидов на виток, еще тоньше, чем В-ДНК и закрученная не вправо, а влево. Торчащие ее на поверхности фосфатные группы образовывали не плавную спираль, а зигзаг, поэтому новый вариант назвали Z-формой.

А-ДНК (слева), B-ДНК (по центру), Z-ДНК (справа)
Это, конечно, не означало, что Уотсон-Криковская модель неверна. Z-форму удалось получить при достаточно экзотических условиях — в растворе с высокой концентрацией солей. И в клетке она тоже получается из В-ДНК лишь при определенных обстоятельствах: например, когда «напряжение» на цепи слишком высоко и его необходимо сбросить. Напряжение появляется из-за чрезмерного скручивания: нити ДНК и так завернуты друг относительно друга, но образованная ими двойная спираль накручивается на какой-нибудь белок (например, гистон), возникает так называемая суперспирализация. Переход в Z-форму помогает сбросить напряжение и развернуть лишние витки — а это, в свою очередь, важно, чтобы с ДНК могли связываться новые белки, например, полимераза при транскрипции.
Поэтому ДНК часто принимает Z-форму при транскрипции генов. Более того, чем больше при этом Z-ДНК, тем активнее идет транскрипция. Гистоны с Z-ДНК связаться не могут, поэтому полимеразе никто не мешает заниматься своим делом. И этим, кстати говоря, активно пользуются опухолевые клетки, у которых левозакрученная спираль вовремя возникает перед нужными им генами.

Башня «Эволюция» (на переднем плане) имеет вид левозакрученной ДНК
Потом нашлись и другие формы двойной спирали. В зависимости от влажности, содержания солей и последовательности нуклеотидов в конкретном участке, ДНК может еще сильнее удлиняться (Е-ДНК) или сжиматься (C- и D-ДНК), включать в себя ионы металлов (М-ДНК) или вытягиваться так, что вместо азотистых оснований в центре спирали оказываются фосфатные группы (S-ДНК). А после того, как в список добавились другие типы внутриклеточной ДНК, вроде ядерной N-ДНК и рекомбинантной R-ДНК (которые, впрочем, попали в этот список не из-за своей формы, а положения в клетке или происхождения), в английском алфавите для вариантов ДНК практически закончились буквы. Тому, кто решит открыть еще какую-нибудь неканоническую форму, придется выбирать из пяти свободных: F, Q, U, V, и Y.
Алфавитный перечень форм ДНК
- A-ДНК — двухцепочечная, чуть толще, чем В.
- B-ДНК — та, которую построили Уотсон и Крик.
- C-ДНК — двухцепочечная, 9,3 нуклеотида на виток.
- D-ДНК — двухцепочечная, узкая: 8 нуклеотидов на виток, содержит много тиминов.
- E-ДНК — двухцепочечная, еще уже: 15 нуклеотидов на два витка.
- G-ДНК — четверная спираль с гуаниновыми тетрадами.
- H-ДНК — тройная спираль.
- I-ДНК — две двойные спирали, которые держатся вместе притяжением своих цитозинов.
- J-ДНК — еще одна тройная спираль, которую образуют повторы АЦ.
- K-ДНК — ДНК трипаносом, особенно богатая аденинами.
- L-ДНК — ДНК, в основе которой лежит L-дезоксирибоза (а не D-, как обычно).
- M-ДНК — В-ДНК в комплексе с двухвалентными металлами.
- N-ДНК — ядерная ДНК.
- O-ДНК — точка начала удвоения ДНК у бактериофага λ.
- P-ДНК — тройная спираль Полинга и Кори.
- R-ДНК — рекомбинатная ДНК (полученная встраиванием чужеродного фрагмента).
- S-ДНК — двухцепочечная, вытянута в 1,6 раз сильнее, чем В-форма.
- T-ДНК — похожа на D-форму, встречается у бактериофага Т2.
- W-ДНК — синоним Z-ДНК.
- X-ДНК — двухцепочечная спираль, которую образуют повторы АТ.
- Z-ДНК — двухцепочечная левозакрученная.
Попасть в переплет
Помимо всевозможных форм двойной спирали и способов ее плетения, ДНК иногда распадается на отдельные нити, которые образуют в шпильки, кресты и другие двуцепочечные фигуры. Случается и так, что уже существующая двойная спираль обрастает новыми соседями.
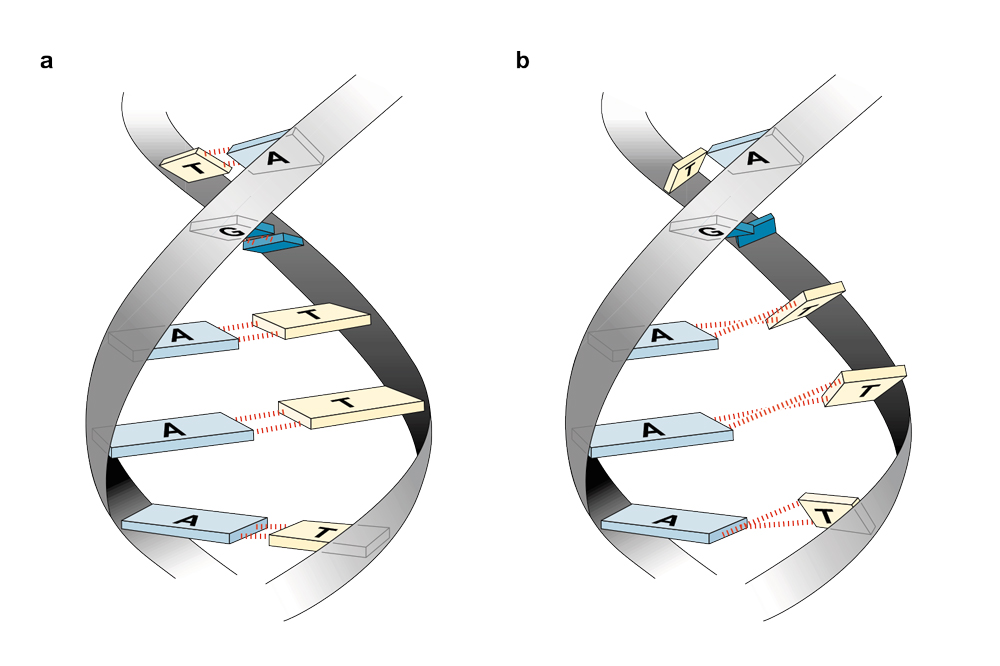
В 1985 году выяснилось, что Полинг и Кори тридцать лет назад были правы: тройная спираль ДНК (H-ДНК) существует. Однако устроена она совсем не так, как они предполагали. В настоящей тройной спирали две цепи соединяются стандартным, Уотсон-Криковским способом, а третья примыкает к ним сбоку, ложась в большую бороздку между цепями. При этом азотистые основания третьей, дополнительной нити соединяются с основными парами не классическим способом, а как бы сбоку — теми самыми связями, которые предсказывал Карст Хугстин. Он тоже, в некотором роде, оказался прав.
Тройная спираль, как и многие альтернативные формы ДНК, тоже возникает в ответ на суперспирализацию цепи. Однако, в отличие от Z-формы, она не поддерживает транскрипцию, а наоборот, ей препятствует. РНК-полимераза, которая привычно расплетает две нити перед собой, не всегда справляется с тем, чтобы разделить триплекс. Поэтому если в гене или его регуляторных областях образуется тройная спираль, он работает хуже прочих.

Варианты образования тройной спирали. Уотсон-Криковские пары обозначены черным, добавочный третий нуклеотид выделен цветом
Бывает и так, что соединяются не две и не три, а сразу четыре цепи ДНК. Чтобы это произошло, в одном месте должны встретиться четыре гуаниновых нуклеотида — и неважно, находятся они на двух цепях одной нити или на четырех разных нитях, не связанных друг с другом. Каждый гуанин образует неклассическую, хугстиновскую пару с двумя соседями, а все вместе они создают квадратную гуаниновую тетраду. Если рядом с ними находятся другие гуанины, способные создать квадрат, то из них складывается стэк — стопка, которая удерживает рядом четыре цепи ДНК.

Гуаниновая тетрада (сверху) и варианты расположения цепей в квадруплексе (снизу)
Все 30 лет, что прошли с момента открытия квадруплексов, количество процессов, в которых они так или иначе замешаны, растет. Известно уже больше двух сотен белков, которые могут избирательно распознавать гуаниновые тетрады — вероятно, последние выполняют роль своего рода генетической разметки, очередного способа регулировать упаковку и транскрипцию генов. Например, они часто встречаются в промоторах (регуляторных участках, с которых начинается транскрипция) разных генов. Совсем недавно ученым даже удалось отличить разные типы рака груди через наборы квадруплексов — от них, в свою очередь, зависело, какие гены в опухолевых клетках были гиперактивны.
Чем дальше мы вглядываемся в молекулу ДНК, тем больше замечаем отклонений от давно привычной модели. Двойная спираль — не единственная и не окончательная структура ДНК, а лишь одна (пусть и самая частая) из поз, которую та принимает в непрерывном танце. Повинуясь велению нуклеотидной последовательности, нить ДНК сжимается и разжимается, изгибается, закручивается и принимает бесконечное число (прекрасных) форм. Ни одна из них — не окончательная: альтернативные структуры ДНК переходят друг в друга, конкурируют с В-формой и между собой, подчиняются сигналам клеточных белков и сами направляют их работу.
Найти и возглавить
Неканонические формы ДНК, при всем своем разнообразии, не возникают в случайных местах. Устойчивость им придает определенный набор нуклеотидов в их составе, поэтому и появляются они лишь в тех участках цепи, где для них есть «удобная» последовательность.
Так, например, в ДНК есть определенные участки, которые особенно охотно сворачиваются в зигзаг. Это места, где чередуются пары Г-Ц: после левого поворота в них каждый второй нуклеотид принимает «неправильную» форму, отсюда и ломаный профиль всей Z-формы. Это означает, что последовательности, склонные принимать Z-форму, можно найти прямо в тексте — если видите ГЦГЦГЦГЦГЦГЦ, то вряд ли прогадаете. Так в одной работе, например, насчитали 391 такой участок в человеческом геноме.
Места, в которых может образоваться тройная спираль, тоже можно узнать по характерной последовательности нуклеотидов. Третья цепь присоединяется либо по принципу комплементарности — то есть к паре Г-Ц добавляется еще один Г, образуя Г-Ц*Г — либо «к своему» — и получается Г*Г-Ц. Поэтому часто такая конструкция возникает в тех местах ДНК, где подряд идет несколько одинаковых (например, ГГГГГ) или химически близких (АГГААГ) нуклеотидов и где они образуют палиндромные (зеркальные) повторы.
Точно также по тексту ДНК можно предсказать и появление квадруплексов. По результатам только одного секвенирования (собственно, прямого перевода ДНК в буквы), в геноме человека их нашлось более 700 тысяч. Не все они, вероятно, встречаются in vivo — для этого соответствующем нитям ДНК нужно оказаться рядом в одной точке сложно устроенного клеточного ядра — однако это может означать, что четырехспиральным структурам отведена какая-то специфическая роль в жизни клетки.
Далеко не всегда образование альтернативных форм ДНК идет клетке на пользу: большинство из них куда менее прочны, чем обычная В-ДНК, и гораздо чаще рвутся. Поэтому последовательности, которые склонны образовывать не-В формы, становятся участками генетической нестабильности и повышенного мутагенеза. Одни исследователи видят в этом двигатель эволюции — если такие участки появляются в генах, связанных с развитием организма. Другие же винят альтернативные формы ДНК во всех видах болезней, связанных со случайными мутациями и перестановками в геноме — от опухолей до шизофрении и аутизма.
Получается, что ДНК содержит не только информацию о строении клеточных белков и РНК, но и о том, какие формы эта информация может принимать, помимо Уотсон-Криковского стандарта. А уже от этих форм, в свою очередь, зависит то, что с этой информацией произойдет: сможет ли клетка ее реализовать или ген, будет вечно молчать, а то и вовсе сломается, породив какие-то дополнительные мутации.
Вероятно, мы научимся однажды вмешиваться в этот процесс — можно было бы, например, построить цепь нуклеотидов, которая имитировала бы третью цепь в спирали и «подсунуть» ее в нужное время в нужном месте, чтобы заблокировать работу какого-нибудь нежелательного гена в клетке. Были даже более смелые предложения — использовать тройную спираль для прицельного редактирования генома: ввести в клетку нуклеотид, который сможет образовать с целевым участком ДНК тройную спираль и побудить систему репарации заменить этот участок на «здоровый» вариант с другой хромосомы.
А пока мы этому только учимся, остается признать структуру ДНК еще одним видом информации — помимо генетической (нуклеотидного «текста») и эпигенетической (доступности генов для считывания) — который несет в себе наш геном. И нам еще предстоит научиться с ним работать, влияя через форму на содержание, или наоборот.
Оригинал earth-chronicles.ru
